Онлайн чтение книги
Рассказ предка. Паломничество к истокам жизни
The Ancestor's Tale: A Pilgrimage to the Dawn of Evolution
Рандеву № 3. Орангутаны
Согласно молекулярным данным, рандеву № 3 (здесь к нам присоединяется орангутан) назначено на 14 млн лет назад – на середину миоценовой эпохи. Хотя тогда на нашей планете уже начало холодать, климат все же был теплее нынешнего, а уровень моря – выше. Из-за этого (а также небольших различий в положении континентов) перешеек между Азией и Африкой, а также значительная часть Юго-Восточной Европы время от времени затапливались. Это важно в контексте размышлений о том, где мог жить сопредок № 3 – наш гипотетический предок в 666666-тысячном поколении. Жил ли он в Африке, как сопредки №№ 1 и 2 – или в Азии? Поскольку он является общим предком человека и азиатских человекообразных обезьян, следует быть готовым к встрече с ним на любом из двух континентов, и тому есть доказательства. В пользу Азии говорит множество окаменелостей, датируемых как раз второй половиной миоцена. С другой стороны, похоже, что именно в Африке появились человекообразные обезьяны – еще до миоценовой эпохи. Различные формы человекообразных обезьян процветали в Африке в раннем миоцене – проконсулиды (несколько видов человекообразных обезьян рода Proconsul) и другие, например афропитек (Afropithecus) и кениапитек ( Kenyapithecus ). Ближайшие современные формы, а также все наши послемиоценовые ископаемые родственники найдены к Африке.
О наших особых отношениях с шимпанзе и гориллами известно лишь несколько десятилетий. Прежде большинство антропологов считало, что человек – это сестринская группа по отношению ко всем человекообразным обезьянам и что он, таким образом, одинаково близок к африканским и к азиатским обезьянам. С общего согласия предпочтение отдали Азии, поскольку именно там жили наши последние миоценовые предки. Некоторые авторитеты даже объявили, что нашли нашего ископаемого “предка” – рамапитека ( Ramapithecus ). Сегодня считается, что рамапитек – то же животное, что и сивапитек (Sivapithecus), – а по законам зоологической номенклатуры приоритет имеет позднейшее название. Таким образом, название “рамапитек” больше использовать нельзя – а жаль, потому что оно прижилось. К сивапитеку (рамапитеку) ученые относятся по-разному, однако многие соглашаются в том, что он близок к линии, от которой произошел орангутан, – а может, он сам непосредственный предок орангутана. Гигантопитек ( Gigantopithecus ) – нечто вроде гигантского наземного сивапитека. Известны и другие азиатские ископаемые того периода. Так, за титул миоценового предка человека борются уранопитек ( Ouranopithecus ) и дриопитек (Dryopithecus). Если бы они жили на “правильном” континенте… Но, как мы увидим, “если” может оказаться реальностью.
Если бы позднемиоценовые человекообразные обезьяны жили в Африке, а не в Азии, в нашем распоряжении имелся бы непрерывный ряд ископаемых, связывающих современных африканских обезьян с раннемиоценовыми формами и богатой африканской фауной проконсулид. После того как с помощью молекулярных данных была абсолютно доказана близость человека к африканским шимпанзе и гориллам, а не к азиатским орангутанам, охотники за предками человека отвернулись от Азии. Ученые решили, что, как бы ни были хороши азиатские человекообразные обезьяны, наша родословная должна относиться к Африке и включать миоценовые формы, – и заключили, что по неизвестным причинам после расцвета миоценовых проконсулид наши африканские предки просто не подвергались фоссилизации.
Это положение вещей сохранялось до 1998 года, когда вышла статья Каро-Бет Стюарт и Тодда Р. Дисотелла “Эволюция приматов – в Африку и из Африки”. В статье (это замечательный пример всестороннего подхода к вопросу) рассказывается о миграциях между Африкой и Азией. И, знаете, сопредок № 3, похоже, жил в Азии. Но об этом после.
А пока задумаемся, как он выглядел. Поскольку сопредок № 3 является общим предком орангутана и современных африканских человекообразных обезьян, он должен быть похожим на любую из них – и одновременно на орангутана. Найдены ли ископаемые, которые могли бы подсказать нам ответ? Что ж, взглянем на генеалогическое древо. К нужному нам периоду относятся люфенгпитек, ореопитек, сивапитек, дриопитек и уранопитек. Наиболее вероятная реконструкция облика сопредка № 3 совместит черты всех пяти азиатских ископаемых родов (при условии, что это животное обитало в Азии). Послушаем “Рассказ Орангутана”, а там решим.
Рассказ Орангутана
Возможно, заявление о наших давних связях с Африкой было поспешным. Что если наши предки покинули Африку около 20 млн лет назад и поселились в Азии, а 10 млн лет назад вернулись в Африку?
Если так, то современные человекообразные обезьяны, включая тех, которые в итоге оказались в Африке, происходят от линии, мигрировавшей из Африки в Азию. Гиббоны и орангутаны – потомки эмигрантов, оставшихся в Азии. Поздние их потомки вернулись в Африку, где раннемиоценовые человекообразные обезьяны уже вымерли, и уже там породили горилл, шимпанзе, бонобо – и нас.
С этим вполне согласуются данные о дрейфе континентов и колебаниях уровня моря. Как раз в нужные моменты в Аравийском море имелись перешейки, соединявшие Азию с Африкой. В пользу этой теории говорит и принцип парсимонии, то есть экономии: хороша та теория, которая многое объясняет, постулируя малое. (Как я не раз говорил, по этому критерию теория естественного отбора Дарвина является, пожалуй, лучшей на свете.) В данном случае речь о том, чтобы до минимума снизить число предполагаемых миграционных событий. С этой точки зрения теория о том, что наши предки все время обитали в Африке (без миграций), на первый взгляд экономнее теории о том, что наши предки сначала переселились из Африки в Азию (первая миграция), а потом вернулись в Африку (вторая миграция).
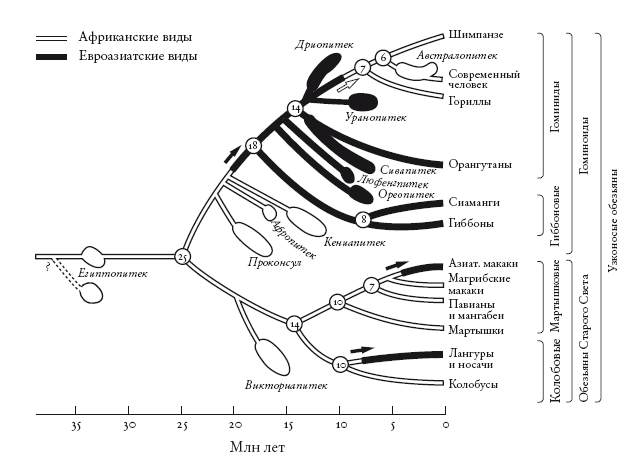
Филогенетическое древо африканских и азиатских обезьян. Расширения обозначают даты, полученные на основе анализа ископаемых, а линии, соединяющие их с древом, построены методом парсимонии. Stewart and Disotell [273].
Но это частный случай: мы рассмотрели лишь собственную линию и проигнорировали остальных человекообразных обезьян, особенно многие ископаемые виды. Стюарт и Дисотелл провели пересчет миграционных событий, в том числе необходимых для объяснения распределения человекообразных обезьян, включая вымерших. Для этого нужно сначала построить древо, отметив на нем все виды, о которых достаточно данных. Следующий шаг – указать для каждого вида, жил он в Африке или в Азии. На диаграмме Стюарта и Дисотелла азиатские ископаемые отмечены черным цветом, африканские – белым. Там представлены не все известные ископаемые, а лишь те, для которых можно определить положение. Стюарт и Дисотелл не забыли и обезьян Старого Света, которые отделились от человекообразных около 25 млн лет назад (самое наглядное различие между ними состоит в том, что первые сохранили хвосты). Миграционные события указаны стрелками.
С учетом этих ископаемых теория переселения в Азию и обратно оказывается экономнее теории о том, что наши предки все время жили в Африке. Оставим хвостатых обезьян, которые, согласно обеим теориям, участвовали в двух волнах миграции из Африки в Азию, и рассмотрим человекообразных. Теория переселения требует всего двух волн миграции:
1. Популяция человекообразных обезьян переселилась из Африки в Азию около 20 млн лет назад. От нее произошли все азиатские человекообразные обезьян, включая современных гиббонов и орангутанов.
2. Популяция человекообразных обезьян вернулась в Африку. От нее произошли современные африканские человекообразные обезьяны, включая нас.
При этом теория постоянного обитания в Африке для объяснения расселения человекообразных обезьян требует шести миграций. И все они направлены из Африки в Азию:
1. Гиббоны (около 18 млн лет назад).
2. Ореопитек (около 16 млн лет назад).
3. Люфенгпитек (около 15 млн лет назад).
4. Сивапитек и орангутан (около 14 млн лет назад).
5. Дриопитек (около 13 млн лет назад).
6. Уранопитек (около 12 млн лет назад).
Разумеется, эти подсчеты верны лишь при условии, что Огюарт и Дис-отелл, опиравшиеся на анатомические данные, верно построили древо. Они, например, считают, что ближайший из всех ископаемых родственник современных африканских человекообразных обезьян – это уранопитек. Его линия ответвляется непосредственно перед линией африканских человекообразных обезьян. Все следующие родственники, согласно оценкам Стюарт и Дисотелла, – азиаты (дриопитек, сивапитек и другие). Если же авторы неверно интерпретировали анатомические данные (например, если ближайший родственник современных африканских человекообразных обезьян – на самом деле ископаемый африканский кениапитек), подсчет миграционных событий надо провести заново.
Сама генеалогическая схема тоже построена по принципу парсимонии. Но мы не пытаемся уменьшить число миграционных событий, необходимых для теории, и игнорируем географию. Вместо этого мы хотим минимизировать число анатомических совпадений (конвергентная эволюция), положенных в основу теории. Так мы получаем древо, не учитывающее географические события, и, чтобы подсчитать миграционные события, накладываем географические данные (черная и белая маркировка на диаграмме). И приходим к выводу: с наибольшей вероятностью современные африканские человекообразные обезьяны – гориллы, шимпанзе и люди – пришли из Азии.
А теперь любопытное замечание. Ричард Клейн из Стэнфордского университета в учебнике по эволюции человека приводит обзор анатомических данных основных ископаемых. Клейн, сравнивая азиатского уранопитека и африканского кениапитека, задается вопросом, кто из них сильнее похож на нашего близкого предка (или родственника) австралопитека. И делает вывод: австралопитек сильнее похож на уранопитека, и если бы уранопитек жил в Африке, он вполне мог быть предком человека. Однако “на основе географических и морфологических данных”, пишет Клейн, кениапитек более подходящий кандидат. Видите? Клейн предполагает, что африканские человекообразные обезьяны просто не могли произойти от азиатского предка, даже если анатомические данные указывают на это. Таким образом, он подсознательно предпочитает географическую парсимонию анатомической. Из соображений анатомической парсимонии уранопитек к нам ближе, чем кениапитек. Однако, хотя об этом не говорится открыто, географическая парсимония берет верх. Стюарт и Дисотелл утверждают, что при рассмотрении географического распространения всех известных ископаемых анатомическая и географическая парсимонии согласуются. То есть географические данные не противоречат утверждению Клейна, на основе анатомических данных признавшему: уранопитек ближе к австралопитеку.
Возможно, этот спор далек от разрешения. Не так-то просто манипулировать анатомической и географической парсимониями. Статья Стюарт и Дисотелла вызвала бурную реакцию: в научных журналах появлялись и восторженные, и разгромные отклики. Мне кажется, что сегодня, оценив все доступные данные, мы должны отдать предпочтение теории “переселения в Азию и обратно”. Два миграционных события – меньше, чем шесть. Кроме того, мне кажется, между позднемиоценовыми азиатскими обезьянами и “нашей” линией африканских обезьян (например австралопитеками и шимпанзе) есть немалое сходство. Таким образом, теория представляет собой компромисс. Исходя из этого, я назначаю рандеву № 3 (и № 4) не в Африке, а в Азии.
Мораль “Рассказа Орангутана” неоднозначна. Когда ученый выбирает между двумя теориями, на первом месте всегда принцип парсимонии. Однако не всегда ясно, как ее оценивать. Наличие хорошей родословной бывает необходимо для дальнейших рассуждений в рамках эволюционной теории. Но чтобы построить хорошее древо, нужен опыт.

ГИББОНЫ. Двенадцать видов гиббонов сейчас объединяют в четыре группы. Порядок ветвления внутри них остается спорным (см. “Рассказ Гиббона”).
На рис. (слева направо): восточный хулок (Hoolock leuconedys), быстрый гиббон (Hylobatesagilis), сиаманг (Symphalangus syndactylus), желтощекий номаск (Nomascus gabriellae).
Нецензурные выражения и дубли удаляются автоматически. Избегайте повторов, наш робот обожает их сжирать. Правила и причины удаления